Notions de Biologie Hyperbare Au cours de la plongée en scaphandre lourd, autonome ou en caisson, nous constatons que le plongeur respire de l'air sous une pression égale
à la pression hydrostatique d'immersion.
à la pression hydrostatique d'immersion.
Son organisme, ses tissus sont constitués par une proportion importante d'eau et de graisse susceptibles de dissoudre les gaz constituant le mélange
respiratoire.
Normalement l'homme vivant à la surface de la terre est saturé en gaz atmosphériques à la pression où il vit.
Le problème qui se pose est de savoir comment les gaz comprimés vont, au cours de la plongée, se comporter vis-à-vis des tissus
Des lois physiques vont encore nous donner l'explication des phénomènes.
A) LOIS DE LA DISSOLUTION DES GAZ
La solubilité d'un gaz dans un liquide, c'est à dire la quantité de gaz dissous par unité de volume d'un liquide est fonction de plusieurs facteurs
1° De la nature du gaz et du liquide.
Autrement dit du coefficient de solubilité du gaz dans le liquide
considéré.
Ce tableau donne la solubilité de quelques gaz dans l'eau
(gaz purs à la pression de 760 mm de Hg).
ml % H2 Hydrogène O2 Oxygène N2 Azote CO Oxyde de carbone CO2 Gaz Carbonique |
à 0° C 2.1 4.9 2.3 3 87.4 |
à 20° C 1.8 5.1 1.6 2.3 17.95 |
L' azote ( N²) est environ deux fois moins soluble que l'Oxygène (0²) et cinq fois moins que le Gaz Carbonique (C0²) dans l'eau (sang), Mais il est beaucoup plus soluble dans les graisses (tissus nerveux, moelle osseuse) que l'oxygène : environ cinq à six fois plus. 2) De la température.
La quantité dissoute avec l'élévation de température, ce que l'on a constaté dans le Tableau ci-dessus. Pour la physiologie humaine, ce facteur intervient peu, la température de l'organisme étant constante autour de 37° C. 3° De la pression.
En effet, la loi de HENRY nous apprend que la quantité d'un gaz dissout dans un liquide à une température donnée est directement proportionnelle à la pression que le gaz exerce sur le liquide. Si l'on porte sur un graphique la quantité de gaz dissout en fonction de la pression, on obtient une droite de pente a correspondant au coefficient de solubilité.
Ainsi du sang placé sous de l'air à des pressions croissantes dissoudra de plus en plus d'oxygène et d'azote.
A l'équilibre, Si nous avons 1,04 ml% de N² à la pression atmosphérique, nous en aurons : 2,08.ml% à 2 Bars,
3,12 ml% à 3 Bars, et ainsi de suite. 4° Du temps.
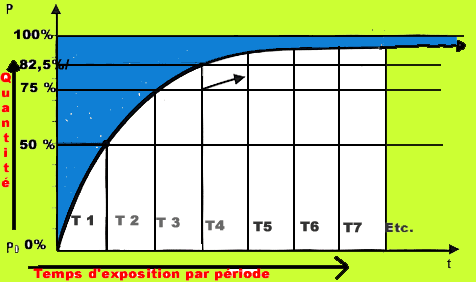
En effet, la dissolution n'est pas un phénomène instantané (pas plus que celle d'un morceau de sucre dans le café) ; elle s'effectue en suivant une courbe exponentielle qui tend vers un équilibre : la saturation. Cette courbe est caractérisée par sa période (T) c'est à dire le temps nécessaire pour atteindre la demi saturation .
Ce phénomène est réversible et si on diminue ou on annule la pression du gaz sur le liquide, le gaz dissout se dégage en suivant une courbe strictement identique mais inversée. Si la variation de pression au dessus du liquide est plus rapide que le rythme du dégagement , le liquide va être sursaturé en gaz par rapport à la pression extérieure. Pour un certain niveau critique de sursaturation, celle ci étant exprimée par le rapport :pression du gaz dissous / pression ambiante, les gaz vont se dégager violemment en formant des bulles. C'est ce que l'on observe lorsque l'on débouche, c'est à dire lorsque l'on met en contact brusquement avec la pression atmosphérique, le
contenu d'une bouteille de champagne ou d'eau gazeuse(Perrier par ex.).
5° Mélange gazeux, Lorsque nous avons affaire, et c'est le cas de l'air, à un mélange gazeux, la dissolution de chacun d'eux est fonction de la pression mais
aussi de leurs concentrations respectives.
Ceci conduit à envisager la notion de pression partielle d'un gaz;
Cette pression partielle d'un gaz (Pp) dans un mélange est donné par la loi de DALTON :
"La Pp d'un gaz dans un mélange est celle qu'aurait ce gaz s'il occupait seul le volume total du mélange et elle est égale à la pression totale du mélange multipliée par le pourcentage de ce gaz dans le mélange".
Donc : Pression partielle = Pression absolue X Concentration.
Ainsi, pour l'air sec à la pression atmosphérique, nous aurons
Pp Oxygène = 1 bar x 21%
= 0, 21 bar = 160 mm de HgPp Azote = 1 bar x 79%
= 0,79 bar = 600 mm de Hg
C'est ainsi que la pression partielle d'azote sera sensiblement la même dans de l'air normal respiré à 20 mètres de profondeur ou dans un mélange à 50%
d'oxygène et 50% d'azote, respiré à 38 mètres de profondeur
Air normal (21/79) à 20 mètres
Pp N² = 3 bars x 0,79% = 2,37 bars
Mélange 50/50 à 38 m
Pp N². = 4,8 bars x 50/ = 2,4
Notez également l'usage normal des symboles pN² , pO², pCO², etc., pour ce qui concerne les pressions partielles.
B) RAPPEL de <<PHYSIOLOGIE de la RESPIRATION >>
Pour comprendre utilement le comportement des gaz respiratoire, il n'est pas inutile d'ouvrir une parenthèse concernant le transport et l'utilisation des gaz respiratoires
Le Tableau suivant donne les valeurs moyennes des pressions partielles et des quantités de gaz dissous ou combinés, dans l'air, le sang et les tissus.
Pression partielle des Gaz en Mm de hg |
O² |
CO² |
N² |
H²O |
Total |
| Air Inspiré
Air Expiré Air Alvéolaire Sang Artériel Sang Veineux Tissus (ou moins ou plus) |
116 100 100 40 30 |
32 40 40 46 50 |
565 573 573 573 573 |
47 (7) 47 47 47 47 |
760 760 760 706 (6) 706 (6) |
Nota 6
La différence avec 760 mm Hg est due aux vitesses de diffusion et de libération des combinaisons chimiques
Nota 7
pH²O ou tension en vapeur d'eau à la température t° de 37° C
Concentration dans les mélanges (Air ou Ml % sang |
O² |
CO² |
N² |
H²O |
| Air Inspiré en %
Air Expiré en % Air Alvéolaire en % Sang Artériel Total Sang Artériel Dissout physiquement Sang artériel Combiné Sang Veineux Total |
20.79
15.26 13.16 20 0.25 9.75 16 |
0.04
4.21 5.26 50 2.69 47.31 60 |
78.42 74.34 75.40 1.04 1.04 0 1.04 |
0.75 6.18 6.18 ------ ------ ------ ------ |
La respiration a pour objet de fournir à l'organisme l' oxygène qui est nécessaire à la combustion du matériel énergétique (fourni par l'alimentation ou ses propres tissus) et à l'évacuation du gaz carbonique produit par ces combustions
L'énergie libérée par ces oxydations est utilisée par l'organisme pour la vie des cellules, le travail musculaire, la lutte contre le froid, etc.
La comparaison des pressions partielles et de concentration en 0² et en C0² de l' air inspiré et expiré montre qu'environ 5 m1 d'0² sont consommés et 4 ml de C0² rejetés pour 100 ml d'air ventilé.
La différence entre l'air inspiré et l'air alvéolaire s'explique par la saturation en vapeur d'eau des alvéoles pulmonaires d'une part et par l'existence d'un espace mort (air résiduel) d'autre part ( voir schéma soufflet)
L'air alvéolaire partiellement renouvelé à l'inspiration est en équilibre avec le sang par l'intermédiaire d'un réseau de capillaires sanguins alvéolaires de très grande surface (100 à 150 m2) .
La totalité du sang de l'organisme (environ 7 litres) met un peu moins d'une minute pour traverser ce réseau..
La succession des phénomènes respiratoires est schématisée ci dessous :
Tableau 6

Le sang veineux amené par les artères pulmonaires, appauvri en 0² et enrichi en C0², arrive au niveau des poumons.
C'est la différence de pression partielle des gaz entre l'air alvéolaire et le sang qui va conditionner les échanges gazeux qui s'effectuent par un simple phénomène de diffusion à travers le mince épithélium des alvéoles.
Donc , p0² alvéolaire étant plus grande (100 mm Hg) que p0² veineuses (40 mm Hg), l'oxygène passe des alvéoles dans 1e sang.
Par contre, pC0² veineuse (46 mm Hg) étant plus grand que pC0² alvéolaire
(40 mm Hg) le C0² passe du sang vers les alvéoles d'où il sera rejeté à l'expiration.
Le sang artériel ainsi renouvelé (hématosé) quitte les poumons par les veines pulmonaires pour faire retour au cur gauche qui va le distribuer par les artères aux différents tissus.
Au niveau des capillaires tissulaires, les phénomènes inverses vont avoir lieu :
0² diffusera dans le sens : sang --> tissus, et le
C0² diffusera dans le sens : tissus --> sang.
Le sang veineux fait retour au cur droit par les veines et le cycle recommence.
L'examen du Tableau ci-dessus et de la figure 6 montre que le sang contient beaucoup plus d'oxygène et de gaz carbonique que ne le laisse prévoir la loi de HENRY.
Ceci est dû au fait que celle-ci ne s'applique qu'à 1a dissolution physique du gaz.
Or, si une certaine quantité, faible d'ailleurs, d'0² et C0² est dissoute à l'état physique, la plus grande partie est sous forme de combinaisons chimiques qui ne suivent plus cette loi, mais sont tout de même sous la dépendance de la pression partielle des gaz.
1° L'oxygène entre en combinaison avec l'hémoglobine (Hb) contenue dans les hématies (globules rouges) pour donner de l'oxyhémoglobine (Hb0²) .
Cette combinaison qui correspond à l'équilibre : Hb + 0² <=> Hb0², suit en fonction des pressions partielles d'oxygène une loi complexe représentée par la figure 7a.
Cette courbe sigmoïde montre pour pO² = 0, toute l'hémoglobine est sous forme réduite (Hb) et que plus la pression partielle augmente plus il y a d'oxyhémoglobine, donc d'oxygène fixé
Toutefois, pour les pressions partielles élevées (voisine de celle de l'air alvéolaire : pO² = 100 mm Hg), la courbe s'aplatit et une augmentation ultérieure de pO² n'assure qu'une faible fixation d'O²
C'est ainsi qu'une hyperventilation permettant d'obtenir une pO² alvéolaire de 120 mm Hg soit 17% d'O² au lieu de la normale qui est de 100 mm Hg soit 13 à 14% d'O², n'augmente le % d'HbO² que de 1 à 2% et cela beaucoup plus du fait de la diminution de la pCO² qui favorise la fixation d'O² sous forme HbO² ( effet Bohr)
L'examen de cette courbe où nous avons noté pour quelques points remarquables la teneur d'oxygène en ml pour 100 ml de sang montre :
Que la quantité maximum d'oxygène fixable sous forme HbO² pour 100 ml de sang est de 20 ml (pouvoir oxyphorique) soit 1/5 ème
Que, à la p0² alvéolaire, le sang artériel : en contient 19,7 ml. Ce fait est intéressant car il montre que la surventilation (9) ou la respiration d'0² pur avant une apnée n'augmente que très faiblement la quantité d'0² fixée: sous forme HbO²
C'est surtout la quantité, dissout à l'état physique qui est dans ces cas augmentée selon la loi de HENRY.
Seul cet 0² dissout est utilisable pour les oxydations cellulaires, l'excès en est donc rapidement utilisé, la dissociation de HbO² n'agissant que pour maintenir une tension d'oxygène dissout compatible avec les échanges cellulaires.
Si donc la pO² augmente fortement dans l'air inspiré du fait de la respiration d'air comprimé ou d'oxygène pur (dans les limites de l'hyperoxie) et par suite la quantité d'0² dissout, l'utilisation rapide de cet oxygène dissout fait qu'il ne jouera aucun rôle direct dans les phénomènes de décompression.
Que, dans les conditions de repos ou de travail modéré, l'organisme n'utilise qu'une faible partie de 1'0² dont il dispose (4 ml %) cette quantité augmente avec l'effort ou dans une apnée par exemple 8 ml% d'0² supplémentaire pouvant être utilisés.
Le reste est inutilisable et ne peut en sortir.
L'oxygène est, lui, livré aux tissus et aux cellules sous forme dissoute et la dissociation de l'oxyhémoglobine assure la constance de cette quantité dissoute, utilisable.
Lorsque la pO² est trop faible, l'oxygène ne peut être utilisé, c'est le cas lorsque le pourcentage d'Hb0² tombe au-dessous de 30 %
2° Le gaz carbonique est en majeure partie transporté sous forme de combinaisons Bicarbonate de sodium (70 %) et composés carbaminés ( Carbhémoglobine 20 %) suivant une loi analogue à celle de l' oxygène.
Il ne faut pas confondre avec Carboxyhémoglobine résultant de la fixation de l'oxyde de carbone sur l'hémoglobine.
Les pourcentages indiqués représentent la contribution des diverses formes dans la quantité de CO² expiré dont les 10 % restant correspondent au CO² dissout
La courbe exprimant la fixation du C0² sanguin en fonction des pressions partielles est une hyperbole équilatère (fig.7 b)
L'examen du Tableau VI montre que le sang contient une quantité importante de C0² sous ferme de bicarbonate (CO 3 Hna), puisque le sang artériel après passage dans les poumons en contient environ 50 ml
Cette quantité fixe constitue ce que l'on appelle la Réserve Alcaline de l'organisme
Outre son importance dans la régulation de l'équilibre acido-basique de l'organisme, ce taux fixe de C0² conditionne la permanence du rythme respiratoire.
En effet, si le CO² est l'excitant normal et le plus important des centres respiratoires bulbaires
L'élévation de son taux dans le sang artériel accélère la respiration (et lorsque ce taux est anormalement élevé., il est la cause de l'essoufflement),
L'abaissement de ce taux ralentit la respiration ou même l'arrête.
Dans l'hyperventilation, ce facteur est un des plus importants pour la prolongation d'une apnée.
Dans l'hyperventilation avec respiration d'un mélange suroxygéné, s'y ajoute le fait que l'augmentation de pO² favorise l'élimination du CO² ( effet Haldane réciproque de l'effet Bohr)
Cette action du C0² nous conduit à rappeler L'élément important de physiologie respiratoire. Les mouvements respiratoires sont sous la dépendance d'un ensemble neuromusculaire comportant
1).Des centres respiratoires (inspiratoires et expiratoires) situés dans le bulbe rachidien, sollicités, c'est à dire excités en permanence et directement par des influences nerveuses et surtout chimiques (pCO² et accessoires p0² du sang artériel qui les irrigue.
2) Des nerfs moteurs partant de ces centres, provoquant la contraction des muscles respiratoires et expiratoires (par exemple muscles intercostaux et diaphragmes).
3) Des nerfs sensibles sollicitant indirectement les centres dont les plus importants sont constitués par les nerfs sensitifs crâniens permettant la parole, le chant, etc., les nerfs venant des chémorécepteurs situés dans le sinus carotidien et l'artère aorte et qui sont sensibles aux variations de p0² (et accessoirement aux variations de pCO²), et des fibres nerveuses du nerf vague provenant de récepteurs pulmonaires sensibles au degré d'inflation et déflation des alvéoles pulmonaires l'inflation donc l'inspiration excitant les centres expiratoires et donc entraînant par voie réflexe l' expiration : réflexe d'Héring-Breuer .
L'énoncé original du réflexe d'Héring-Breuer ( 1868 ) disait que : " l'inspiration appelle l'expiration et vice versa".
Confirmé pour l'hyperpnée mais pas pour l'eupnée, les phénomènes étant plus complexes.
Si l'alternance rythmique des mouvements inspiratoires et expiratoires est sous la dépendance des nerfs sensibles des poumons, le fonctionnement constant des Centres respiratoires est surtout sollicité par la pCO² (directement) et la p0² (réflexe des chémorécepteurs).-
Le second facteur est surtout important en anoxie (diminution du taux d'O² dans l'air inspiré) et le facteur normalement dominant est le premier, c'est à dire la pC0²..
On a pu dire : « que le gaz carbonique était une véritable hormone respiratoire rég1ant son propre taux physiologique » Toute élévation dans le sang artériel entraînant une hyperventilation ( éliminant le C0² en excès) et toute diminution, une hypoventilation (donc retenant le CO² )
3° Pour fermer cette longue parenthèse, il nous reste à considérer le cas du gaz inerte : l'azote (ou dans les mélanges artificiels utilisés en plongée : l'hélium , l'hydrogène par exemple), diluant l'oxygène atmosphérique (ou du mélange) ,
Comme le montre le tableau VI, son taux est constant dans le sang artériel et le sang veineux, ce qui montre qu'il n'est pas consommé par l'organisme. Son transport obéit uniquement à des lois physiques de dissolution. Malheureusement, en ce qui concerne la plongée ou la respiration d'air comprimé, sa teneur élevée (79% de l'air) pose de grave problème de sa dissolution progressive et de son évacuation à la remontée.-
C'est ce que nous allons envisager dans la suite de ce chapitre.
C. - CONSÉQUENCE DE LA DISSOLUTION DES GAZ
Pendant la plongée ou la vie sous pression les gaz vont se dissoudre (selon les lois vues précédemment : pression et temps) dans le sang qui se sature rapidement grâce à l'énorme surface de contact que représentent les fins capillaires des alvéoles pulmonaires..
Le sang artériel ainsi saturé arrive aux différents tissus et leur livre des quantités de gaz proportionnelles à l'écart entre la pression des gaz sanguins et des gaz tissulaires.
Cet écart diminue au fur et à mesure que les tissus se chargent et la dissolution rapide au début, tend ensuite lentement vers la saturation..
Des tissus les gaz viennent diffuser de proche en proche vers les tissus voisins.
Cette dissolution sera naturellement fonction de la vitesse de circulation et de l'irrigation plus ou moins grande des différents territoires tissulaires.
Elle tend vers l'état d'équilibre qui est la saturation.
Lors de la remontée, donc au retour vers la pression atmosphérique, ces phénomènes vont jouer en sens inverse dès que la pression des gaz intrapulmonaires sera inférieure à celle du sang veineux arrivant aux poumons.
Les tissus seront alors en état de sursaturation.
Deux cas peuvent alors se présenter :
1) Si la remontée est lente ou plus précisément s'effectue dans les conditions prévues par les tables de plongées les gaz dissous vont se libérer des poumons et ceux des tissus diffuseront dans le sang, jusqu'à atteindre leur taux normal de saturation à la pression atmosphérique.
2) Si la remontée est trop rapide, l'état instable de sursaturation, défini plus haut, atteint sa valeur critique et les gaz se dégageront dans les tissus eux mêmes sous forme de bulles pouvant être entraînées dans la circulation et créer des obstacles empêchant des échanges vitaux.
Ces bulles sont responsables des "accidents de décompression" connus sous diverses dénominations . "Mal des caissons", "Aéroembolisme", "Coup de pression", "Maladies des scaphandriers".
Suivant la localisation et l'importance des bulles les accidents seront plus ou moins graves et se manifesteront de diverses manières
* Accidents cutanés (puces, moutons),
* Ostéomusculaires, bends
* Nerveux (paralysie, paraplégies, etc. ...Ces problèmes seront traités en détail par ailleurs.
La prévention de ces accidents repose sur le respect des tables de décompression donnant pour chaque profondeur d'immersion et durée de séjour : les vitesses de remontée, les durées des paliers près de la surface et les coefficients de plongées successives.
Le but des paliers de décompression est de maintenir le coefficient de sursaturation au-dessous de sa valeur critique par une élimination progressive des gaz dissous durant la descente et le séjour au fond.
Leur traitement s'effectue par recompression en caissons et décompression lente après soulagement des douleurs ou disparition des troubles.
D.PRINCIPES DU CALCUL D'UNE TABLE DE PLONGÉE
Les données ci-dessous sont issues des travaux publiés par le pharmacien-chimiste en Chef du G.E.R.S. PERRIMOND-TRONCHET en 1962.
Les bases théoriques de ce calcul sont des notions de physique et de physiologie indiquées au cours de ce chapitre. Nous n'avons toutefois considéré dans le cycle respiratoire que trois éléments :
Air alvéolaire,
Sang,
Tissus.
En ce qui concerne les tissus il y a lieu de préciser que dans le cas du calcul d'une table il faut considérer l'organisme, comme divisé en divers tissus, différents par leur composition chimique et par leur vascularisation (irrigation sanguine plus ou moins abondante et rapide
On peut les caractériser anatomiquement (tissus sanguins, musculaires, nerveux adipeux, osseux) et en fonction de leur vitesse de saturation en gaz par leur période T définie plus haut.
C'est ainsi que l'on peut définir des tissus se saturant très rapidement comme le sang (période de 3 à 4 minutes), des tissus lents comme la moelle épinière et les tissus adipeux (période supérieure à 120 minutes) et des tissus intermédiaires.
Pour des raisons de simplification on choisit par exemple trois tissus de période « T »
T = 40', 75' et 120',
en adoptant pour chacun un coefficient de sécurité (correspondant à un coefficient de sursaturation admissible c'est à dire inférieur à la valeur critique) qui est par exemple respectivement de 2,3 pour le tissu 40' et de 2 pour les tissus 75' et 120'.
Connaissant la durée du séjour au fond (la durée de la descente étant incluse dans ce temps) des courbes du type de la. fig.5b ou leurs abaques linéaires correspondantes permettent de déterminer le pourcentage de variation de tension (ou pression) des gaz dans les divers tissus.
En multipliant cette valeur par la différence de pression positive ou négative
( 1 ) on obtient la valeur absolue de cette variation et en y ajoutant la pression initiale ( 2 ), on obtient la pression finale des gaz dissous pour le temps et la profondeur donnée.
( 1 ) Les Lois de saturation à la descente, et désaturation à la remontée, sont admises comme étant symétrique mais de signe opposé.
( 2 ) Tous les tissus, au départ , sont supposés en équilibre avec la pression atmosphérique
Cette valeur divisée par le coefficient de sécurité donnera pour chaque tissu la profondeur du premier palier.
On choisira naturellement la profondeur la plus grande (en général imposée par le tissu le plus rapide).
La durée de séjour à ce palier sera conditionnée par la tension de gaz admissible à atteindre pour permettre la remontée au palier suivant (ou à la surface s'il n'y a qu'un seul palier à 3 mètres
Cette tension est égale au produit :
Pression hydrostatique du palier suivant X coefficient de sécurité et, la durée sera celle nécessaire pour que la tension finale définie plus haut tombe à cette valeur.
Elle sera donnée par la courbe ou l'abaque linéaire;
Donc on calcule le % de variation entre :
Tension finale / Tension admissible en fin de palier et on lit le temps correspondant pour le tissu considéré
S'il y a plusieurs paliers, on refait les mêmes opérations
Le but des paliers ainsi calculés est de maintenir la tension des gaz dissous au-dessous de la valeur critique et non d'assurer une désaturation complète de l'organisme.
Celle ci est en effet, surtout vers la fin, en raison de la forme exponentielle du phénomène, très lente.
Si bien que durant plusieurs heures après la remontre, l'organisme contient une quantité de gaz dissous supérieure à la normale et cela surtout dans les tissus de période longue, dont la désaturation comme la saturation est la plus lente.
Ces tables tiennent compte de cela et le coefficient C de plongée successive indique précisément la tension de gaz dissous dans le tissu le plus lent choisi, au moment de la sortie de l'eau.
Ce coefficient impose une majoration `a ajouter à la durée réelle d'une plongée ultérieure pour rentrer dans la table.
Cette majoration diminue lorsque augmente l'intervalle entre les plongées et s'annule lorsque cet intervalle dépasse 6 heures alors que la désaturation est pratiquement totale pour tous les tissus
Enfin, il faut insister sur le fait qu'une table est calculée en fonction d'une vitesse de remontée donnée (20 mètres/Minute pour la table du Ministère du Travail 1990), le temps de remontée est alors négligé dans les calculs.
Si la remontée s'effectue à une vitesse plus lente les tissus lents dont la tension finale est plus faible continuent à se charger entre le fond et la profondeur du palier.
Un cas particulier est posé pour la plongée en altitude, la pression atmosphérique y étant inférieure à celle du niveau de la mer et par suite pour une même profondeur, la différence de pression entre la surface et le fond sera plus grande en altitude.
Cette différence intervenant dans le calcul des tables vu précédemment, il importe donc de calculer une profondeur et une pression fictive en mer correspondantes.
La formule calculée par le G.E.R.S. donne cette profondeur
P = p H/h
<p> étant la profondeur réelle atteinte en lac,
<H> et <h> les pressions atmosphériques respectivement au niveau de la mer et au niveau du lac de montagne, exprimées en mètres d'eau ou en mm de Hg (10 m = 760 mm Hg).
Ainsi à 2.000 mètres une plongée à 40 mètres correspondra à P = 40 X 760/ 595 = 50 mètresC'est donc avec cette profondeur de 50 mètres qu'il faudra entrer dans la table et le palier correspondant à 3 mètres en mer devra se faire à
3 = (p) X (60/595) d'ou p = 3 X 535/760 = 2,34m.
E)..En guise de conclusion à ce chapitre il convient d'ouvrir une dernière parenthèse,
Les tables de plongées ne peuvent avoir une valeur absolue.
Leur calcul repose sur des données où un certain empirisme règne encore en 2019, leur expérimentation n'est que statistique et par suite leur application vaut pour la moyenne des individus d'un type moyen et d'état physiologique normal (ou peut être plus simplement courant), et pour une plongée standard.
La dispersion des individus, c'est à dire l'écart à cette moyenne explique parfaitement que certains ayant respecté une table peuvent malgré cela avoir un accident et que d'autres ne les ayant pas respectées s'en tirent indemnes.
Juger sur ces cas extrêmes n'est pas logique, c'est sur l'ensemble d'une population que doit porter ce jugement.
Lorsqu'un individu ne rentre plus dans le cadre moyen, sommairement défini plus haut (corpulence particulière, travaux sous-marins, dans des conditions particulières de t°, d'efforts, etc. ) il est bien évident que les précautions doivent
être renforcées, les coefficients de sécurité augmentés et le choix d'une table de décompression plus sévère envisagé .
De même, si l'on plonge loin de toute possibilité de traitement, il y a intérêt à se placer dans les conditions les plus favorables et donc choisir la table couvrant la sécurité la plus grande.
=============
Pour ce qui nous concerne, nous considérons que les tables Ministère du Travail, sont plus appropriée aux activités des Scaphandriers Amateurs que celle du Ministère des Armées cette dernière étant adaptée sur un standard de Militaire de Carrière , jeune, super entraîné et constamment en forme « olympique » ce que n'est habituellement pas le cas du Plongeur de loisir ou Scaphandrier Amateur.
==============
Le choix des tables reste possible, les tables françaises Marine Nationale, Ministère du Travail à l'air jusqu'à 60 mètres et aux mélanges au-delà , les tables américaines, les tables anglaises, italienne et suisse
Point n'est besoin de se quereller, le choix est libre et facile.
Si de la discussion jaillit la lumière, par contre querelles et polémiques sont stériles et au contraire vont à l'encontre du but recherché en semant le trouble et la confusion chez ceux dont les connaissances ne leur permettent pas de se faire une opinion personnelle.
Le but est d'appliquer une table en sachant les raisons qui commandent son utilisation et les bases qui ont servi à son établissement
Le rôle du Scaphandrier Instructeur est d'assurer l'enseignement, nécessaire pour atteindre ce résultat.
page precedente dernière modification : Jeudi, 24-Jan-2019 15:00